The
Physiology of Transport Substances in the Blood (Sodium)
By Professor Marcel Uluitu, M.D. Ph.D.
Co-Authored by Diana Popa (Uluitu), M.D.
Department of Microbiology, Immunology
and Molecular Genetics
[Editor’s Note: This paper is presented as Part V of a
series of chapters from the new book “The Physiology of Transport Substances in
the Blood (Sodium)”; preceeding and subsequent chapters have been/will be
featured in recent past/upcoming issues of this Journal. This segment features
Chapter Six].
Chapter 6
Physiology and Physiopathology of Sodium
Transport in the Blood
I have presented
above the mechanisms through which the transport of Na+ in
the blood is made, from which the idea
derived that the normal functioning and normal excitability are compatible with
low chemical activity of Na+ within the method described above (218,
232, 233). In other words, Na+ is transported in a state of
interaction with proteins. The process has been described in humans (Figure 21
and Table no.21.) and rats (figure 20 and table no. 23), in normal
collectivities in the case of both species, this is a dominant distribution of
about 75%.
Figure 21. The
number of anionic sites (![]() M 5HT) of heparin in school children in a normal collectivity
(236).
M 5HT) of heparin in school children in a normal collectivity
(236).
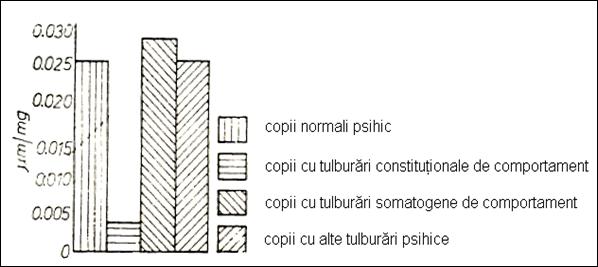
Table No. 21.
Detailed table with children of the two lots which include mechanisms for
transport of Na in the blood (236)
|
No. |
Name |
Personal antecedents which may influence the brain |
Dificultăţi familiale |
Type A questionnaire Attitude towards : |
Woodworth-Matthews test |
Psychiatric conclusions |
Activity of
serum cations += free (not bound - = bound |
|||||||||
|
Educational excess |
Disintegrated families |
Lack of
supervision |
School |
Teachers |
Own self |
Family |
Colleagues |
balanced |
Tendencies |
|
Degree of instability |
|||||
|
|
LOT I
1. |
|||||||||||||||
|
1. |
B.D. |
- |
- |
- |
- |
0 |
0 |
0 |
0 |
0 |
+ |
- |
Mentally normal |
- |
- |
|
|
2. |
B.C. |
- |
- |
- |
- |
0 |
0 |
0 |
7 |
7 |
+ |
- |
Mentally normal |
- |
- |
|
|
3. |
B.D. |
- |
- |
+ |
- |
0 |
0 |
0 |
0 |
0 |
- |
High emotivity |
Mentally normal |
- |
- |
|
|
4. |
B.G. |
- |
- |
- |
- |
0 |
0 |
0 |
0 |
0 |
+ |
- |
Mentally normal |
- |
- |
|
|
5. |
B.D. |
- |
- |
- |
- |
0 |
0 |
0 |
0 |
0 |
+ |
- |
Mentally normal |
- |
- |
|
|
6. |
C.G. |
- |
- |
+ |
- |
0 |
0 |
0 |
0 |
0 |
+ |
- |
Mentally normal |
- |
- |
|
|
7. |
C.G. |
- |
- |
- |
- |
0 |
0 |
0 |
0 |
0 |
- |
Slightly impulsive and epileptical |
Mentally normal |
- |
- |
|
|
8. |
C.F. |
- |
+ |
- |
- |
0 |
0 |
0 |
0 |
0 |
+ |
- |
|
- |
- |
|
|
9. |
C.C. |
- |
- |
+ |
- |
0 |
0 |
7 |
7 |
0 |
+ |
- |
|
- |
- |
|
|
10. |
C.R. |
- |
- |
- |
- |
0 |
7 |
0 |
6 |
0 |
+ |
- |
|
- |
- |
|
|
11. |
D.A. |
- |
- |
- |
- |
0 |
0 |
0 |
0 |
0 |
+ |
- |
- |
- |
- |
|
|
12. |
D.G. |
- |
- |
- |
+ |
0 |
0 |
0 |
0 |
0 |
- |
Depressive and
hypochondrical |
- |
- |
- |
|
|
13. |
D.D. |
- |
- |
- |
- |
0 |
0 |
0 |
0 |
0 |
- |
Very slight emotivity |
- |
- |
- |
|
|
14. |
G.A. |
- |
- |
- |
- |
0 |
8 |
6 |
8 |
6 |
+ |
- |
Mentally normal |
- |
- |
|
|
15. |
I.D. |
- |
- |
- |
- |
0 |
0 |
0 |
0 |
0 |
- |
Intense emotivity , depressive and hypochondrical |
Mentally normal |
- |
- |
|
|
16. |
M.F. |
- |
- |
- |
- |
0 |
0 |
7 |
7 |
0 |
+ |
- |
Mentally normal |
- |
- |
|
|
17. |
N.B. |
- |
- |
- |
- |
0 |
0 |
10 |
9 |
0 |
- |
Very slight emotivity |
Mentally normal |
- |
- |
|
|
18. |
P.G. |
- |
- |
- |
- |
7 |
7 |
10 |
11 |
0 |
+ |
- |
Mentally normal |
- |
- |
|
|
19. |
P.G. |
- |
- |
- |
- |
0 |
0 |
0 |
0 |
0 |
+ |
- |
Mentally normal |
- |
- |
|
|
20. |
R.A. |
- |
- |
+ |
- |
- |
- |
- |
- |
- |
+ |
- |
Mentally normal |
- |
- |
|
|
21. |
T.I. |
- |
- |
- |
- |
0 |
0 |
0 |
0 |
0 |
+ |
- |
Mentally normal |
- |
- |
|
|
|
2. “OTHER PSYCHIC DISTURBANCES” |
|||||||||||||||
|
22. |
B.C. |
- |
+ |
- |
- |
- |
- |
- |
- |
- |
+ |
- |
Inhibiory type pubertary disturbances |
1 |
- |
|
|
23. |
C.A |
- |
- |
- |
+ |
11 |
11 |
8 |
9 |
0 |
- |
Very slight emotivity |
Mixed behavioral disturbances |
1 |
- |
|
|
24. |
D.N. |
- |
- |
+ |
- |
- |
- |
- |
- |
- |
- |
Slightly schizoid , paranoic and depressive |
Reactive depression in involution |
0/1 |
- |
|
|
25. |
F.A. |
- |
- |
+ |
- |
0 |
0 |
0 |
9 |
0 |
+ |
- |
Purbertary behavioral disturbances |
|
|
|
|
26. |
G.M. |
- |
- |
- |
- |
0 |
10 |
14 |
12 |
0 |
- |
Shlightly depressive , hypochondrical and unstable |
Essential pubertary and reactive behavior |
2 |
- |
|
|
27. |
M.M. |
- |
- |
- |
+ |
- |
- |
- |
- |
- |
+ |
- |
Slight pubertay disturbances due to social reasons |
1 |
- |
|
|
28. |
M.M. |
- |
- |
+ |
- |
8 |
8 |
8 |
9 |
8 |
- |
Obsessive |
Essential instability with pubertary opposition
elements |
1/2 |
- |
|
|
29. |
M.D. |
- |
+ |
- |
- |
- |
- |
- |
- |
- |
+ |
- |
Slight behavioral disturbances , instability ,
reactive elements |
1 |
- |
|
|
30. |
T.D. |
- |
- |
+ |
- |
- |
- |
- |
- |
- |
- |
Intensely reactive |
Reactive behavioral disturbances |
0/1 |
- |
|
|
|
3. TRAUMATIC |
|||||||||||||||
|
31. |
M.A. |
Trauma at birth . Reanimation with O2 |
- |
- |
- |
0 |
7 |
11 |
11 |
9 |
- |
Paranoic and schizoidal |
Slight reactive depression. Microeffects |
0/1 |
- |
|
|
32. |
M.P. |
Effects of meningo-encephalitis |
- |
+ |
- |
8 |
7 |
12 |
18 |
9 |
+ |
- |
Slight instability. Organic behavioral disturbances |
1 |
- |
|
|
33. |
N.A. |
Forceps. Eye bleeding |
- |
- |
- |
- |
- |
- |
- |
- |
- |
Neurotic |
Uşoară instabilitate. Dezinteres
şcolar |
0/1 |
- |
|
|
34. |
T.N. |
Front-nasal trauma |
- |
- |
+ |
7 |
12 |
9 |
15 |
7 |
- |
High instability , and slightly emotive, depressive
and hypochondrical |
Tulburări mixte de comportament esenţiale
şi reactive |
2 |
- |
|
|
|
|
|||||||||||||||
|
35. |
A.D. |
- |
- |
- |
- |
0 |
9 |
0 |
0 |
8 |
- |
Emotivity combined with a slight depression |
Essential instability. Choreic movements of fingers |
2 |
+ |
|
|
36. |
C.D. |
- |
- |
- |
- |
0 |
0 |
0 |
0 |
0 |
+ |
- |
Behavioral and personality disturbances |
1/2 |
+ |
|
|
37. |
D.A. |
- |
- |
+ |
- |
- |
- |
- |
- |
- |
- |
Instability, emotivity |
- |
- |
+ |
|
|
38. |
G.C. |
- |
- |
+ |
- |
0 |
0 |
6 |
17 |
|
- |
Slight emotivity |
Slight essential disturbances , anxiety , emotions |
0/1 |
+ |
|
|
39. |
G.M. |
Brain traumatism with unconsciousness |
- |
- |
- |
0 |
7 |
10 |
8 |
0 |
- |
Sligh paranoic and schizoidal forms |
Mixed essential behavioral disturbances and lesional
instability |
2 |
+ |
|
|
40. |
G.T. |
- |
- |
+ |
- |
- |
- |
- |
- |
- |
- |
Slight instability and emotivity |
- |
- |
+ |
|
|
41. |
S.M. |
- |
- |
+ |
- |
- |
- |
- |
- |
- |
- |
Instability |
Essential reactive disturbances |
2 |
+ |
|
|
42. |
S.M. |
- |
- |
+ |
- |
0 |
12 |
9 |
15 |
0 |
- |
Depressive, hypochondrical and instability |
Essential and reactive behavioral disturbances |
2 |
+ |
|
6.1.
Determining the health status in humans
Determining the health status in humans was carried out with means that are
refer to the physical and mental status. It is necessary to detect subclinical
manifestations as well , when normal school collectivities are studied(1).
6.1.1.
General health status.
This was investigated by clinical examinations on medical instruments,
laboratory tests, medical records at the school medical rooms, psychological
examinations, psychic pathology etc.
6.1.2.
The mental health status.
The investigations on Mental Health made use of multiple methods for showing
some known factors involved in the origin
of psychic disorders (19).
6.1.2.1. Personal history.
Prenatal and postnatal periods.
6.1.2.2. Heredo - collateral history.
6.1.2.3. Socio – familiar background .
Educational opportunities in the family, family conditions, population density
per room.
6.1.2.4. Preschool period. The appreciation of parents.
Evaluation of parents on : psychomotor development up to school age, evaluation
of parents on behaviour in the family, the quantity and quality of sleep,
phobias, parents’ opinion about child’s
friends, relations with parents.
6.1.2.5. Investigations at school .
Educational results , opinion of
teachers about the child, relations with
the teachers, complex process of behavioral integration.
6.1.2.6. Clinical psychiatric examination.
Clinical examination and psychiatric test for assessing the balance between the
processes of excitation and inhibition.
6.2.
Determining the chemical activity of blood sodium.
6.3.
The classification of subjects.
Results of tests presented above (Table 21) on 44 subjects of 12 years for both
sexes show a group (1) of 8 children with high chemical activity, it is homogeneous,
all having constitutional type behavioral disorders.
The second batch in which the chemical activity of Na+
in the blood is very small being screened through interactions with proteins is composed of
psychically normal subjects : children with neuropsychic disorders with brain
lesion history. A last group is formed
of children with various psychiatric disorders on puberty background without behavioral deviations.
The formation of the two lots, with Na+ transported in the ionic
state with high chemical activity and a second group to which the chemical
activity of Na+ is unsignificant (141, 60) was also made at rats , by selection of animals from
normal collectivity , estimating the
response to acoustic stimulation (109, 132, 19, 59, 192, 60).
6.4.
Non excitabile function structures and mechanisms of transport of blood Na.
An important parameter is to compare the response of non excitable cells to
those excitable at physiological
stimuli. Non excitabile cells have also polarized membranes (MP) by the
participation of potential K+, but without AP. At their polarization
, Na+ has a contribution of 1%. Below there is a comparision of parameters of non excitable organs in
subject groups with sodium ionic activity to those where Na+
linked to proteins is inactive.
6.4.1. Total proteins and Na (flamfotometric) in the blood.
The values of total blood sodium (flamfotometric) and plasma protein levels
(Table no. 22) shows no differences between the two groups: (I) without any
chemical activity of blood Na+, (II) with transport in the ionic
state.
Table
22. Biological blood constants in humans in relation to the mechanism of
transport of Na (224)
leucocytes hemoglb red cell group. hematocrit blood
glucose. protein mEq of sodium Na
group RBC WBC
hgl hcr gly prot
Na mEq
mil mii
(I)
4,187 6.311 13,33![]() 35.72
35.72![]() 0.87
0.87![]() 5.7
5.7![]() 151.1
151.1![]()
0,13 0.33 0.01 0.15
1.27
(II)
4.05 6.750 12 56![]()
![]() 34, 05
34, 05![]() 0 9
0 9![]() 6.04
6.04![]() 151.1
151.1![]() . 0.43 1.05 0.03
0.3 1.16
. 0.43 1.05 0.03
0.3 1.16
6.4.2. Respiratory function.
It is illustrated by red blood cell number, hematocrit, globular values
(tabel.22), chest development (27), spirometry (25). These do not indicate
differences between lots formed on the basis of transport mechanisms of Na.
Both groups present values of respiratory function corresponding to age (Table
23).
Table
23 Spirometric static and dynamic data in
group I (Na in interaction with proteins) and II (Na transported as
ions) (224).
Group Vt
F VE VO
CV CV% VEMS
VEMS% VIMS
I![]() SD 592 18,4
10 310 2958
2199 2617 89
2778
SD 592 18,4
10 310 2958
2199 2617 89
2778
28 07
05 10 95
33 72 1
92
II![]() SD 541,6 21.4
11 301 2798
2194 2515 90
2614
SD 541,6 21.4
11 301 2798
2194 2515 90
2614
45.8 1.7
0.8 10.8 193 59.4
154,4 1 6 175
6.4.3.White blood cells.
Similar situation is found on line leucocyte (Table No 22 )
6.4.4. Carbohydrate and lipid
metabolism (224, 100).
Glucose values (Table No.22) of insulinemy and response after a standard effort
(Table no. 24) - test of pancreas
reactivity - shows that the transport mechanisms of Na are not important for regulating carbohydrate
metabolism for both groups (224), which range within normal values (138). There
have been established no differences in terms of triglycerides, cholesterol,
phospholipids and serum lipase in children with psychiatric disorders (100).
6.4.5.
Growth and Development ( Somatometric
values , GH ).
Somatic parameters analysis values (Table nr.25), the level and reactivity to
an effort to standard growth hormone (Table nr.26) shows that between these two
groups there are no quantitative differences in the development of these
functions , as there is no excitable
tissue in the sense discussed above.
Table 25 Comparison of
somatometric indices of children in group I
(Na without chemical activity, with group II (children
with abnormal behavior of constitutional type ) (224)
circumferences
diameter
Group weight
height pubis head chest abd
pelvis biacr bithr
I 39.5
151 80 53.3
71.8 61 78.6
33.1 26.6
M![]() SD 1 1.3
0.8 0.2 0.8
0.7 0.9 0.4
0.1
SD 1 1.3
0.8 0.2 0.8
0.7 0.9 0.4
0.1
II 38
150.7 80.2 54
67.5 64.1 75.4
33.3 26.7
M![]() 2.1 2.4
1.7 0.4 3.3
3.0 3.7 0.8
1.1
2.1 2.4
1.7 0.4 3.3
3.0 3.7 0.8
1.1
But the release of GH response to the test ladder
requires a more detailed analysis that does not have in view the processes of
Na transport in the blood but are present in children with organic injuries of
the nervous system (Table nr.26), in history (36). In this case, low reactivity
of GH (104, 105, 130, 162) in the ladder test can not be attributed to
disturbance of the mechanisms of Na interaction - compounds polianionic plasma,
which otherwise are not even present - but rather recognize postlesional
effects.
Table
26: The GH blood concentrarion in children with normal transport of Na (Group
I) and those with high activity chemical Na (Group II) and response to the test
ladder (224)
before the effort after
effort
unlesional 6 9 0 7 11
6 1 1 P 0, 01
I
I
lesional 4 2 0 9 7
03 1, 07 P = insignificant.
II unlesional
6 75 0, 81 10
05 1 34 P 0, 02
before the effort after effort
unlesional M![]() SD 6.9 11 6 P
SD 6.9 11 6 P![]() o.o1 Grouo
I 0.7 ].]
o.o1 Grouo
I 0.7 ].]
lesional M![]() SD 4 2 7 03 P=ansignif ] 1.09 1.07
SD 4 2 7 03 P=ansignif ] 1.09 1.07
Unlesional GroupII M![]() SD 6 75 10 .05 P
SD 6 75 10 .05 P![]() 0.01
0.01
0.81 1.34
In conclusion ,
the function of tissue and non excitable organs presented here is not
correlated with the transport of Na in the blood - the interaction with
proteins or ionic state - its role is associated with general metabolic
processes which have been discussed.
6.5. The function of the excitable structures and Na transport in the
blood of humans.
The functional level of excitable tissue has been measured at the same two groups for both
species. (Table No. 22, 25, 26) (Figure No. 21 and table No. 21): (1) subjects
in which Na is transported by intreaction with blood proteins (without chemical
activity) and (2) Na having chemical activity on the couple reactance heparin /
serotonin.A problem raised is represented by the need to determine
parameters of excitability in both species . In humans , the great
variety of living and genetic
conditions , (127) influences the
development , while the somato-psychic state of health requires verification of the components of
the studied batch with the means mentioned (26), and other methods to provide
additionally data regarding excitability. From the examination of table
no.21 it results that the group of
examined children contains two subgroups. A subgroup includes children who have
Na with significant chemical activity, which accompanies abnormal behavior
constituti (1) some intensity clinic, which accompanies constitutional behavior
disorders , some of clinical intensity , which are accompanied by educational
disorders and by unbalance of inhibition processes and excitation. Another
subgroup, where no chemical activity of serum Na is detected ,
is non homogenous. Some of them present postlesional psychic disturbances (95,
96). A second subgroup with abnormal type prepubertary disorders, but not
behavioral, and a third group of mentally normal children. These data are
supplemented by the resuls of EEG examinations
, psychic aptitudes , etc.and establishment of some neuro-endocrine mechanisms
where Na is involved to maintain the homeostasis of circulation.
6.5.1.
Electroencephalography activity.
EEG serves as the most direct indicator of the state
of vigilance and the cortico-cerebral excitability (45, 63, 78, 240). The
analysis of cortico-cerebral electrical signals (45, 64, 105, 130) requires
processing of signals obtained from recording selected according to intentional
local activation , or in case of clinical investigations, imposed by the need
of diagnostics, etc. Brain excitability investigation in children from the mentioned groups was made by one single parieto - occipital derivation ,(163)
under standard conditions: (63 240) with
open eyes in mental relaxation or in a mental effort (solving a test of
attention), followed by activation of ![]() rate at the closing of eyes (fig.22). The records are
postprandial and before conducting the school. EEG activity in humans (163,
164) shows variations in the frequency and amplitude as described. EEG signals
are processed (164) as the F / V (frequency / voltage) (Figure 22, 23 and Table
No. 27) which allows a statistical analysis and insurance between different
epochs in the same subject or identical parts from different subjects:
bioelectric activity in children with Na in the interaction with serum proteins
and children with Na transport in ionic state.
rate at the closing of eyes (fig.22). The records are
postprandial and before conducting the school. EEG activity in humans (163,
164) shows variations in the frequency and amplitude as described. EEG signals
are processed (164) as the F / V (frequency / voltage) (Figure 22, 23 and Table
No. 27) which allows a statistical analysis and insurance between different
epochs in the same subject or identical parts from different subjects:
bioelectric activity in children with Na in the interaction with serum proteins
and children with Na transport in ionic state.
Ratio (F / V) (64, 240) is an integrative size of EEG
segments. Its value is a sensitive indicator for brain activity in humans (163,
164, 240, 241). High values of the ratio
(F / V) is observed when on the EEG route there prevails the ![]() rate . Alteration
of values F / V is an important
indicator of the overall level of activation (240). The ratio value allows
comparison with the basic status of tensional status . From the statistical
analysis of EEG data obtained from children (Table No. 27) and (Figures 22, 23)
show that the "normallly psychic" children with normal behavior in
which transport of Na in the blood is in a bound form , the values of the (F / V) ratio decrease significantly upon the
closing of eyes, while children with Na+ wth chemical activity in the blood, with constitutional disturbance
behavior record a deficit . The installation
of rhythm
rate . Alteration
of values F / V is an important
indicator of the overall level of activation (240). The ratio value allows
comparison with the basic status of tensional status . From the statistical
analysis of EEG data obtained from children (Table No. 27) and (Figures 22, 23)
show that the "normallly psychic" children with normal behavior in
which transport of Na in the blood is in a bound form , the values of the (F / V) ratio decrease significantly upon the
closing of eyes, while children with Na+ wth chemical activity in the blood, with constitutional disturbance
behavior record a deficit . The installation
of rhythm ![]() (Table no.27, Figure No. 23), describes the deficit for these
subjects (140). The deficit is not found in the other two subgroups of
children: postlesional and endocrino -
puberty (Table No. 27). It is to be noticed that this deficit is to be found
particularly in children with somatogenic abnormal behavior where Na+ is
screened in the blood (104, 105, 130, 162) .
This calls for a regulating role transport for Na+ transport in the blood, in interaction with proteins,
on brain excitability, the bound transport of Na+ being compatible with the normal excitability
of the brain. The increased chemical activity of Na+ blood is accompanied by nervous
hyperexcitability. Thus the response of the body to the sensory stimulation can
advocate for the phyisiology of the blood transporter of Na+.
(Table no.27, Figure No. 23), describes the deficit for these
subjects (140). The deficit is not found in the other two subgroups of
children: postlesional and endocrino -
puberty (Table No. 27). It is to be noticed that this deficit is to be found
particularly in children with somatogenic abnormal behavior where Na+ is
screened in the blood (104, 105, 130, 162) .
This calls for a regulating role transport for Na+ transport in the blood, in interaction with proteins,
on brain excitability, the bound transport of Na+ being compatible with the normal excitability
of the brain. The increased chemical activity of Na+ blood is accompanied by nervous
hyperexcitability. Thus the response of the body to the sensory stimulation can
advocate for the phyisiology of the blood transporter of Na+.
Table no.
27 The ratio F / V of the EEG in different conditions in children studied (236)
|
|
OE (ochi deschişi,
prima secvenţă) |
|
CE (ochi închişi, secvenţa a 2-a) |
OE-CE |
TEST (efort psihic de atenţie concentrată) |
|
CEt (ochi închişi după efort psihic) |
TEST- CEt |
|
NORMALI |
444.13±18.30 |
P<0.05 |
263.40±24.12 |
180.70±14.70 |
541.50±17.00 |
P<0.05 |
263.30±17.90 |
212.50±17.90 |
|
CONSTITUŢIONALI |
422.50±34.40 |
P<0.01 |
308.80±54.50 |
113.70±29.50 |
416.70±33.50 |
P<0.01 |
284.80±33.60 |
131.80±10.20 |
|
Semnificaţiile faţă de normali |
|
|
|
P<0.05 |
|
|
|
P<0.02 |
|
LEZIONALI |
433.00±35.40 |
P=N |
347.50±80.00 |
85.50±51.60 |
449.25±24.20 |
P=N |
316.00±75.20 |
133.25±69.20 |
|
Semnificaţiile faţă de normali |
|
|
|
P<0.05 |
|
|
|
P=N |
|
ALTE TULBURĂRI PSIHICE |
398.30±13.50 |
P<0.01 |
231.30±13.30 |
167.00±19.80 |
424.90±27.00 |
P<0.01 |
237.00±13.10 |
185.00±28.30 |
|
Semnificaţiile faţă de normali |
|
|
|
P=N |
|
|
|
P=N |
Figure
22.Cerebral bioelectric activity in a 12-years child with (1) eyes open and (2) with eyes closed
(236)
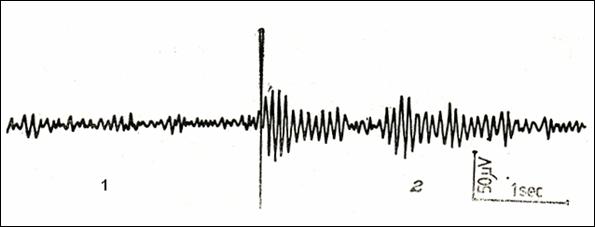
Figure 23.
Variation in the ratio F / V (frequency / amplitude) of EEG in a normal psychic
subject with Na transport in interaction with serum proteins. 1 = eyes open
mind at rest; 2 = eyes closed; 3 = eyes open, effort to resolve a test of
attention, 4 = eyes closed. The immediate installation of rhythm α
upon reopening the eyes is to be noticed (236)
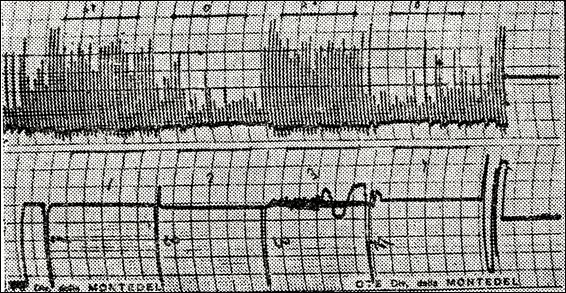
Figure 24
Changes in the F / V (frequency / amplitude) of EEG in a child with hyperactive syndrome with seric Na transport in a
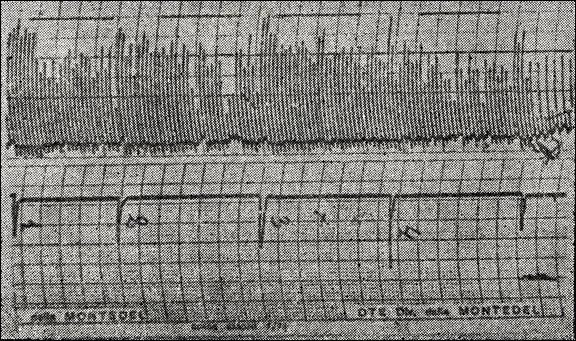
6.5.2.
Psychic funtions.
Determining the mental aptitudes, in centiles, in
conjunction with Na+ seric
activity suggests the influence the
latter has on intrinsic cerebral excitability on socio-intellectual and
adjustive performances . Importance has been given to the processes of
concentrated and distributive (90) attention, to imeediate auditive memory for
words (24), psycho-neurotic tendencies
(90), a type A questionnaire for
determining the overall image of the child adapteded to the population of
6.5.2.1.
Concentrated attention.
In children
with disturbances of constitutional type
behavior concentrated attention is affected , as well as in children with behavior of somatogen type disturbances (1) , thus the
targeting attention deficit compared with mentally healthy subjects with
blood Na screened is showing a deficit .The disturbance of concentrated
attention is accompanied by hyperactivity. As regards the quantity of concentrated attention, this is only
showing deficit in children with somatogenic behavior (95, 96, 104, 105) , with neuronal
affections, inexistent in children with constitutional behavior disturbances.
6.5.2.2.
Distributive attention.
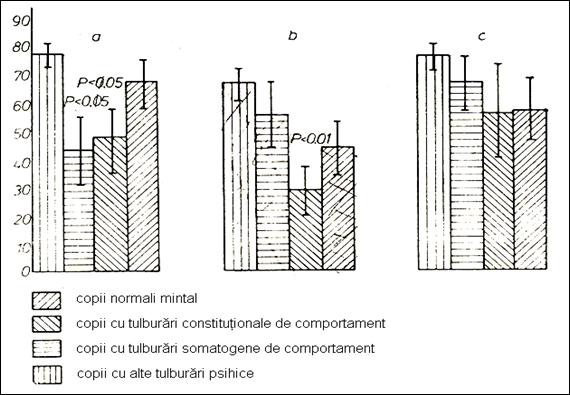
The total distributive attention shows a deficit only
in children with somatogenic disturbances (Figure no. 25). Children with behavioral disorders of
constitutional type present a very good distributive attention.
Figure 25. The quality and quantity of attention in
children investigated in relation to the mechanism of transport of serum Na
(236)
6.5.2.3. Memory.
Immediate auditory memory and intelligence have
complex producing mechanisms ,
insufficiently known. They are dependent on the processes of attention
and on the neuronal capacitaty to store information and especially the
ability to extract and to reproduce (1). In this context, mentally normal
children , with mechanisms for the transport of Na+ normal, have the
coefficients of memory close to 100, while the other categories of groups are
located below the value of 50 centiles.
6.5.2.4.
Intelligence.
Intelligence does not show differences between groups
formed by the mechanisms of transport of Na in the blood.
6.5.3.
Cardiohemodynamic function.
6.5.3.1.
General
Distribution of mass blood in the vascular bed is
determined by the occasional functional level of certain organs and apparatus:
postprandial digestion activity, physical effort of various types and
intensities, the body position (clino-ortostatism), the action of external
factors on the body (positive acceleration), mode of exposure to heat sources,
etc.
In these conditions, the maintenance of circulatory homeostasis takes place
through keeping an optimum ratio between the blood volume of vessels and that
which is provided by the adequate irrigation of each tissue.
Adjusting mechanisms are multiple and complex, nervous
and humoral. The dissociation of these mechanisms and evaluation of every one
of them is difficult, the most relevant parameters being the cardiovascular
hemodynamics and hydroelectrolitics, but the most important being the
sodium.From among cardiohemodynamic parameters,the most representative are:
heart frequency, systolic and diastolic blood pressure during clinostatism and
after passing to ortostatism, pulse pressure, index of ventricular work (Figure
26).
Figure 26. Cardio-vascular parameters in
clino and ortostatism in groups I and II
of children mentioned (224).

6.5.3.2. Mechanisms for regulating cardiac and vascular functions.
The response capacity of the heart and vessels to
action of disturbing factors also depends on the nature of transport of Na+ in the
blood. The parameters mentioned show differences between subjects where serum
Na transport is in the form of binding comparatively to those in which it is
transported in the ionic state (Table No. 21, Figure 21). The existence of free
ionic Na+ in serum is accompanied by lower values of the parameters
mentioned in clinostatism (noted "0" in fig 26, 27).
By passing to passive ortostatism by means of an
automatic tilting table there is induced an accumulation of blood in the lower
region of the body,thus producing variations of parameters recorded minute by
minute, for 10 minutes, to get the full picture of the effort of adjusting.
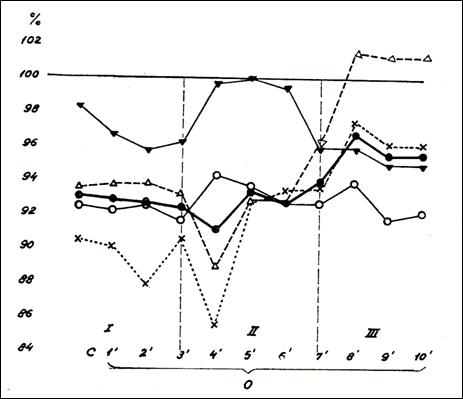
Figure
27. Cardio-vascular parameters normalized, during the changing posture of the
groups I and II (224)
6.5.3.3. Compensation of hemodynamics.
The regulation of hemodynamics during posture change
to ortostatism has two phases: first phase of six minutes from posture change
includes rapid intervention mechanisms of the autonomic sympathetic nervous
system to modify the diameter vessels, followed by the shifting of blood toward
the heart (224): a second phase is slow, with long duration and is intensified
by the humoral system renin-angiotensin-aldosterone-sodium, and natriuretic
polypeptides with action on vascular muscles. Their release depends on Na+
action at kidney level. In addition, retention of Na under endocrine control,
results in the restoration of mass blood
circulating. This phase begins after the six minutes change of posture, in
ortostatism.
6.5.3.4.
The evolution of hemodynamic parameters after the change of posture (Fig. 26,
27)
This period highlights the differences between the two
groups of children, formed on the basis of different mechanisms of the
transport of blood Na+ with reference to the regulation of
cardiohemodynamic function. (fig, 26, table No. 25). In the group containing
the subjects having serum Na transport in the ionic state there is noted a trend for maintaining the distribution of the blood through an
increased mass heart rate , with arterial blood pressure showing a deficit in
the first six minutes of ortostatism. In minutes 9-10 the deficit also invoves
the diastolic pressure, expressing the disturbances in the humoral
renin-angiotensin-aldosterone-Na mechanism
and in the natriuretic peptides (148).
The picture obtained is similar to that induced by the
Na+ loading of the organism (230, 231, 232, 236). Moreover, research
on humans in outer space has demonstrated that
compensation of hemodynamics and of psychiatric disorders can be
obtained by the administration of water and Na+ (38, 76, 187, 247.
148). The addition of hormons, steroids and antidiuretics substantially
restores hemodynamic parameters (76, 146, 186).
6.6. Excitable systems function in rats.
6.6.1. The influence of hypersaline regimen on
electrocorticography (ECG).
In rats there has been studied (223) the influence of
hypersaline chronic regimen compared to a group maintained at hyposaline
regimen, by recording bioelectric cerebrocortical activity . The two groups
were kept, from intrauterine life until the age of 6-8 months with a different
intake of NaCl: both groups received an identical diet, normal, but one group
received for drinking physiological
solution ( hipersaline ) and another group received distilled water. In both
groups there was recorded the electrocorticography. There were used silver
electrodes , chronically implanted on the cortex, in the parieto-occipital region(223). Animals
prepared in this way were placed in individual
soundproof cages , obscured,but
having a 6-W bulb mounted for
light stimulation (197)or for recording in a light-up conditions.
Electrocorticography processing by determining the F / V (164)ratio is
inadequate, even though upon a simple
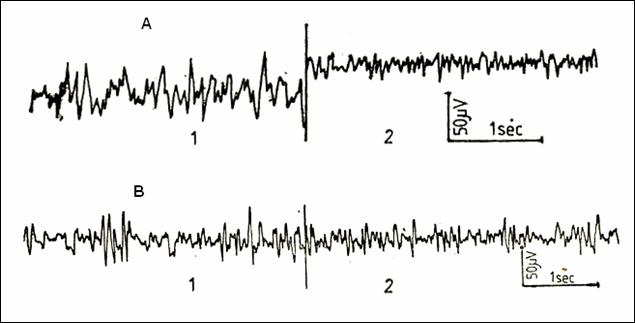
inspection clear changes are noticeable (Figure 28). Probably, in rats,
sensory stimulation induces conjugated variations of ECG potentials in frequency
and amplitude, resulting in a constant value of the ratio F / V. The ECG
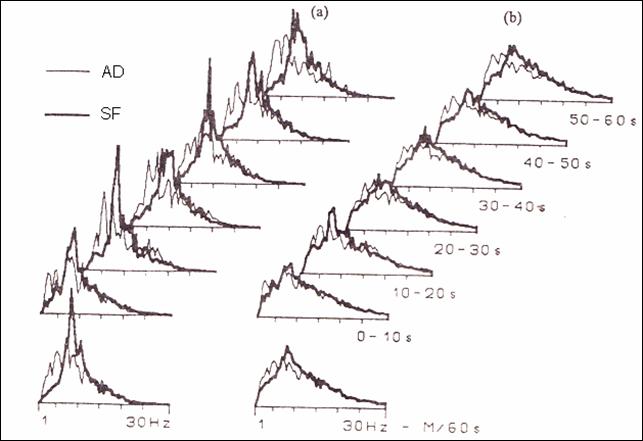
processing in this case is done by applying Fourier transformant (223, 237),
with general applicability (figure 29) .
Figure
28.The electrocorticographic activity
(ECG) in rats: A = chronically hypersaline regimen; B = chronically
hyposaline regimen; 1 = reference, recording in the dark; 2 = stimulation by
noise on dark background (223)
Figure
29. Comparing average ECG of rats consuming chronic hypersaline solution with the rats having a hyposaline one (223)
(Fourier transformant).
6.6.2.The chemical activity of blood sodium,
correlated with the neuromuscular excitability in rats.
The auditory stimulation is the preferred method to form the groups of rats by selection of
animals susceptible to audiogenic convulsions. For this purpose , every single
animal is placed in a cage (236) covered with a transparent lid through which
is monitored rat behavior during auditory stimulation. The noise is generated
by an electric bell placed under the cage.
Most of the animals (225, 117, 229) remain calm, or
even inspect the enclosure, to establish
the origin of the noise. Other animals are hypersensitive, expressed by
hypermotricity, a state of panic, convulsions, opisthotonus followed by a state
of weariness and sometimes death.
Determinig the chemical activity of Na+ in blood shows its absence
in normal animals, nonsensitive to noise, but it is present, in accordance with
the intensity of functional manifestations, in hyperexcitable animals (Table No.
28).
Table nr28. Variation of the number of anionic sites
of heparin, as HT ![]() g / 1 mg heparin, in the presence of rats blood serum with
various types of response to acustic stimulation (227)
g / 1 mg heparin, in the presence of rats blood serum with
various types of response to acustic stimulation (227)
behavior ![]() g 5HT/mg heparin
g 5HT/mg heparin
calm
3 1![]() 1,04
1,04
agitated 2,74![]() 1,69 P = 0, 52
1,69 P = 0, 52
convulsions 2,14![]() 0,72 P = 0, 024
0,72 P = 0, 024
agitated +
convulsions 2,42![]() 1, 26 P = 0, 086
1, 26 P = 0, 086
The absence of chemical activity of Nations or its
reduction is therefore compatible with calm behavior during acoustic
stimulation in rats.
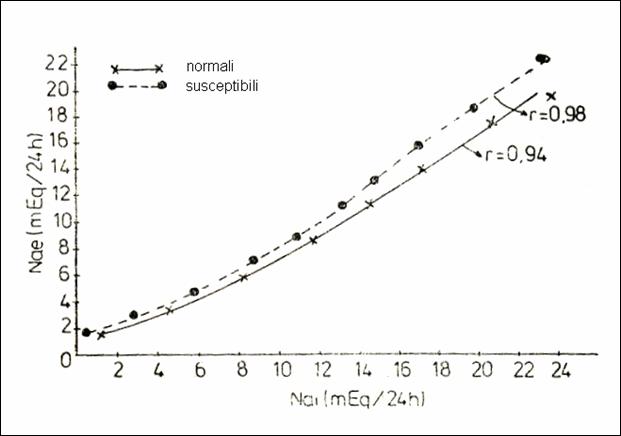
Figure 30. Elimination of Na according to the ingestion (mEq / 24h)of sodium
in normal rats and in rats susceptible to acoustic convulsions (225)
6.6.3.
Motor activity.
Animals susceptible to audiogenic seizures are hyperactive not only during acoustic
stimulation, but they have also free behavior in the cage (No table. 29) or
motivated behavior (table No. 30)
Table 29.Total motor activity of
the two groups recorded during free behavior in the cage. (227, 230,
229, 223)
references (with Na bound), calm during stimulation) 353.87![]() 149.5
149.5
animals hypersensitive to noise (Na in the ionic state, convulsions) 517.41![]() 264.8
264.8
P = 0 .001.
It can be noticed that the animals in which blood
serum Na+ is transported freely present a significantly more intense
activity than normally excitable animals, in agreement with the behavior
induced by acoustic stimulation.
Table
No. 30. Motivated motric activity (for distilled water and physiological
solution)at their free choice,in groups of animals with normal excitability and
hyperexcitable animals (227)
steps to: distilled water physiological solution
normoexcitable 191,61![]() 86,15 162,25
86,15 162,25![]() 80,57
hyperexcitable 266,56
80,57
hyperexcitable 266,56![]() 146,19 250,85
146,19 250,85![]() 136,92
136,92
p![]() 0,01
p
0,01
p![]()
From the tables
it results that the intensity of normal motor behavior is compatible only when
blood Na+ is transported in
interaction with proteins, without chemical activity for anionic grups of
heparin, while the chemical activity of cation is accompanied by
hyperexcitability, expressed by intense motor activity, comparable to that
induced by a chronic hypoproteic regimen (192, 193, 194).
6.6.4. Hemodynamic compensation.
Rats exposed to disturbances of hemodynamics
through hypergravitation (angular
acceleration = G + 5) (76) in centrifuges for small animals, repeatedly , (224)
is followed by changes of hemodynamic and Na+ homeostasis. In rats
with normal excitability to noise and blood transport of Na+ in the
interaction with proteins, the centrifugation induces the decrease of
elimination of renal Na+ (148) from the ingested fraction (Figure
no. 31). In rats whose Na+ is transported in ionic state, with
susceptibility to audiogenic seizures
(228) , exposure to hipergravitation increase is followed by renal loss
of sodium, which expresses an inadaptable reaction (Fig. 31) (229, 186, 149).
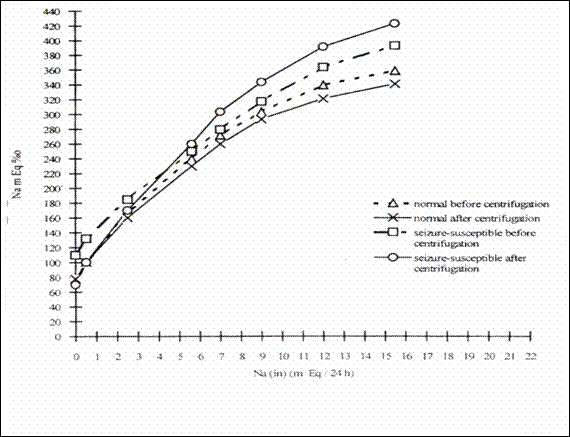
Figure 31 The influence of repeated exposure to
hypergravitation (+5 G) upon the elimination of renal Na in the normoexcitable
rats and in rats , susceptible to audiogenic seizures and with Na in the ionic
state in the blood. (Δ-normal before centrifugation;-x – normal after
centrifugation, ¤ - rats susceptible to convulsions before centrifugation; - ○ susceptible to convulsions after
centrifugation ) (229).
6.6.5.
Mineralocorticoid function in connexion with the transport mechanisms of blood
Na and of age.
Investigation of the mineralocortcoid function is also
done by determining a synthetic indicator
of the function, the value of the ratio urinary Na / K. Researches conducted on
the same two groups, normals adults , and another group of animals susceptible
to audiogene convulsions (Tab No. 26 and
Fig. 32) (230, 229, 223, 227).
The ratio of urinary Na and K concentration (Na / K) is different in the two
groups (Fig 32). The value is significantly higher in animals with chemical
activity of Na+, expressing a functional lower level of the
mineralocorticoid system (Fig.32). The ratio Na / K is significantly different
in these animals and after repeated centrifugation (+5G for 30 min x 5 times),
while in the normoexcitable animals the value of ratio is decreasing. It
expresses the differences of the systems involved in regulating the balance of
the increased hydrosalin (78). There has been noted differences between the two groups regarding
the mineralocorticoid activity (Fig 33) (230). In adults, the value of the
ratio Na / K is significantly higher in animals susceptible to audiogene convulsions , having therefore less mineralocorticoid activity than
normoexcitable animals. The differences between the two groups is emphasized
with age. In normal animals, age induces diminution of the mineralocorticoid
activity (Fig 33), also known (63, 212, 199, 67, 241), due to senility process
of involution (5, 190, 199, 212), between 17 and 27 months age and decreased
secretion of aldosterone. In the animal with Na+ sanguin transported
in active chemical state , the ratio Na / K decreases (Fig 33) to values
comparable to those in the
normoexcitable adult animal, which suggests an improvement in
mineralocorticoid activity. A picture in the mirror over the normoexcitable
animal is carried out (Fig 33). This evolution of mineralocorticoid activity
can recognize different causes.
Reduction of the concentration capacity of Na+ at the kidney level
(Fig.33) having as a result the increase of the circulant Na+ and
the decrease of K+ elimination, that stimulates aldosterone
secretion (78,105,216). Another mechanism could recognize an up-regulation
process maintained by the low
concentration level of aldosterone in the blood (63) in this group of animals. Genetic factors could also be
incriminated (190) in the carrier protein synthesis, but it is known that
protein malnutrition is accompanied by increased content of corticosteron in
the blood in rats (129, 189) concomitant
with the increase of the fighting
behavior.
Figure 32. Variation of the ratio urinary Na / K acc. to mEq Na+ ingested by normoexcitable
animals and by those with Na+ transport in ionic state (227)
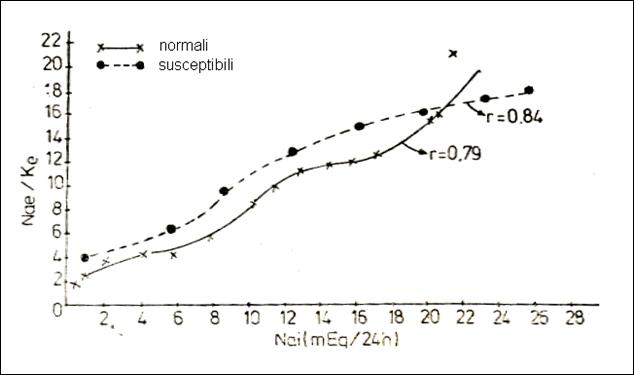
Figure 33. Hydroelectrolitic metabolism in rats
related to age and to the mechanisms of
Na+ transport in the blood serum (230).
6.6.6.
Hydroelectrolytic balance as referred to the Na transport mechanisms.
Analysis of brain activity decodification also includes the estimation of
hydroelectrolytic homeostasis in the two groups of rats: normal, with absent
chemical activity of Na , and the group susceptible to audiogenous convulsions
with Na+ transport in ionic state. Hypersensible animals consume
significantly more saline solution than
normal animals (Table no. 31) in freely motivated behavior, than distilled
water. (227)
Table 31.
Consumption of distilled water (ml/24 hours) and saline solution (ml/24 hours)
in the two groups of rats (227) :
groups distilled water saline solution P(same group)
reference 12,43![]() 6,75
10,76
6,75
10,76![]() 8,56 P
8,56 P![]() 0,18
0,18
convulsion 11,39![]() 5,82
15,85
5,82
15,85![]() 9,31 P
9,31 P![]() 0,126
0,126
P(between groups) ![]() 0,57
0,57 ![]() 0,034
0,034
The animals with Na+ ionic state transport
consume more liquids than the normal ones (Table 32)
Table 32. Total liquids (distilled water + saline) in
normal and spasmodic rats (ml/24 hours) (220).
groups (
ml/24 h)
reference 24,03![]() 9,3
9,3
hypersensitive 27,23![]() 6,9
6,9
P
0,063
The same picture is presented by the renal elimination
of Na+ (Figure 30) (242). Hyperexcitable animals with Na+
transport in ionic state lose a larger amount of Na+ corresponding
to that ingested as intrinsic motivation
, on choice. Data presented in tables 31 and 32 show that in both groups of
animals there is no motivation for
liquid in free access conditions to both vessels. However, animals susceptible
to audiogenous convulsions, with higher
Na+ serum activity than the reference system show
significantly increased preference for the solution of NaCl as compared with
normal animals, as well as for the distilled water vessel , thus compensating
the mentioned renal losses (Fig. 30). In
these animals there is an unbalanced matabolism of sodium by which motivational
processes are stimulated as well.
[The Final Summary and the Bibliography
will be featured in the upcoming
May-June 2010 issue of this Journal.]
Professor Marcel Uluitu, M.D. Ph.D. began his
scientific activity in Physiology in 1953 at the
Professor Uluitu has also investigated
cerebral tissue excitability, studying the structure modification of the
protein macromolecules, and the physiological and pathopysiological processes
in which are involved Sodium and Lithium. He implemented an original method for
physical and chemical processes which involve the chemic active sodium, in
normal processes and in the cerebral excitability dysfunctions, in human and in
experimental model (animal). These results of this work gave him the chance to
outline the chapter herein relating to the physiology of substances transport
in the blood. This is based on the physical and chemical interaction between
blood components.
His papers are included in the
collections of the U.S. National Library of Medicine and the U.S. National
Institute of Health. He is a member of the
Dr. Diana Popa (Uluitu) is a
researcher in the Department of Microbiology, Immunology and Molecular Genetics
at the